
Abstract
Dans l’enseignement on définit souvent les classes par la présence d’écailles notamment pour les poissons. Or des chercheurs de l’Université de Genève ont établi récemment comment ces phanères (couverture tégumentaires) ont pu évoluer au cours des temps en des formes aussi diverses que des écailles, des plaques osseuses ou une peau nue, mettant à mal les critères de classification souvent utilisés en classe.
Les poissons à nageoires rayonnées (actinoptérygiens) constituent le groupe de vertébrés le plus diversifié au monde. La majorité de ces poissons possèdent des écailles comme protection couvrant leur corps. Cependant, plusieurs lignées présentent une armure corporelle composée de plaques osseuses ou sont dépourvues de toute structure de protection. La diversité et les transitions entre les différents types de couverture de tégument n’avaient pas été étudiées auparavant dans un cadre évolutif. Pour étudier comment cette peau a évolué au fil du temps Lemopoulos & Montoya‐Burgos, (2021), ont reconstitué – à partir de nouvelles phylogénies basées sur la comparaison de séquences d’ADN – l’évolution des structures protectrices de la peau chez les poissons, remontant jusqu’à l’ancêtre commun de tous les poissons à nageoires rayonnées, il y a plus de 420 millions d’années.
Ils ont montré que seuls des poissons qui avaient perdu leurs écailles ont ensuite pu développer une armure osseuse et que le type de protection de leur peau influençait leur choix d’habitats en eau libre ou sur le fond de la mer. Cette étude, publiée dans la revue Evolution Letters, fournit une nouvelle explication de l’incroyable diversité de cette lignée de poissons, qui comprend plus de 25 000 espèces.
Jump-To-Science discutera les implications pédagogiques. Juan Montoya‐Burgos a accepté de fournir à Jump-To-Science (JTS) pour ses lecteurs des séquences permettant d’effectuer avec les élèves des alignements pour comprendre la manière dont on établit le degré de similitude entre divers espèces de nos jours. Cf plus bas une activité en classe élaborée avec cet auteur, et Marie-Claude Blatter du SIB.
 encourage le lecteur à aller vérifier dans l’article d’origine : ici
encourage le lecteur à aller vérifier dans l’article d’origine : ici
![Fig 1: Les poissons à nageoires rayonnées (actinoptétygiens) ont souvent des écailles (Squalius cephalus), ont parfois la peu nue (Ictalurus punctatus) ou munie de plaques osseuses (Pterygoplichthys multiradiatus) [img]. Source :Press release UNIGE](https://www.unige.ch/communication/communiques/files/3616/1589/9563/page_garde_Montoya.png)
Fig 1: Les poissons à nageoires rayonnées (actinoptétygiens) ont souvent des écailles (Squalius cephalus), ont parfois la peu nue (Ictalurus punctatus) ou munie de plaques osseuses (Pterygoplichthys multiradiatus) [img]. Source :Press release UNIGE
Résumé de l’impact (article d’origine commenté par JTS)
Les chercheurs étudient quelle protection tégumentaire était présente à l’origine des poissons à nageoires rayonnées et comment de nouvelles structures protectrices sont apparues et ont évolué au fil du temps.
Les chercheurs montrent qu’un tégument écaillé était le plus répandu durant l’évolution des poissons à nageoires rayonnées, mais que la perte des écailles s’est produite plusieurs fois de manière indépendante, alors que l’acquisition d’écailles à nouveau ne s’est presque jamais produite.
Cela peut être un exemple pour montrer que même les modèles simples de l’évolution vus en classe peuvent expliquer que les mutations produisent plus fréquemment des pertes de structure ou de fonction.
De plus, ils montrent que les téguments sans écailles ont très probablement conduit ces espèces à modifier leur écologie et à coloniser le plancher des océans et des plans d’eau.
Les modèles simples de l’évolution vus en classe expliquent bien que parmi les variants sans écailles celles vivant dans des milieux différents mais favorisant ces variants ont pu se reproduire avec un plus grand succès que la plupart des autres. On peut imaginer des exercices ou des questions d’évaluation en les basant sur un article scientifique récent…
Les avantages fonctionnels d’un tégument sans écailles en milieu benthique restent à démontrer, mais l’augmentation de la respiration cutanée pourrait être une explication.
On peut aider les élèves à utiliser ces modèles pour montrer que si ces variants sans écailles ont pu se reproduire avec succès dans un milieu différent comme sur le fonds des mers et des lacs c’est qu’elles avaient acquis de nouvelles capacités pour mieux vivre, se nourrir et se protéger des maladies dans ce nouveau milieu, et donc la concurrence des espèces à écailles n’y était pas très forte.
Les auteurs montrent que les plaques osseuses du tronc ont également émergé de manière indépendante plusieurs fois au cours de l’évolution des poissons à nageoires rayonnées, mais ces armures protégeant le tronc ne semblent apparaître qu’après un événement de perte des écailles. Par conséquent, si les acquisitions de plaques osseuses du tronc sont phylogénétiquement indépendantes, elles ont besoin d’un «même terreau» pour émerger.
Pris ensemble, leurs résultats prouvent que les structures tégumentaires ont contribué à la diversification exceptionnelle des poissons à nageoires rayonnées Lemopoulos, A., & Montoya‐Burgos, J. I. (2021). Traduction automatique révisée ( les remarques en vert sont de Jump-To-Science)
encourage le lecteur à aller vérifier dans l’article d’origine : ici
Les méthodes qui ont permis d’établir ces conclusions
![Fig 2: Reconstruction de traits ancestraux par cartographie stochastique de la présence / absence d'écailles sur un arbre phylogénétique de 11638 espèces d'Actinopterygii. Les clades rouges indiquent des taxons écaillés, tandis que la couleur bleue correspond à des taxons sans écailles. Les images correspondent aux espèces représentant des lignées présentant du TBP. [img]. Source : Lemopoulos & Montoya‐Burgos (2021).](https://onlinelibrary.wiley.com/cms/asset/3a6d07ad-d1c3-4712-ac03-6f961af78a73/evl3219-fig-0002-m.png)
Fig 2: Reconstruction de traits ancestraux par cartographie stochastique de la présence / absence d’écailles sur un arbre phylogénétique de 11638 espèces d’Actinopterygii. Les clades rouges indiquent des taxons écaillés, tandis que la couleur bleue correspond à des taxons sans écailles. Les images correspondent aux espèces représentant des lignées présentant du TBP. [img]. Source : Lemopoulos & Montoya‐Burgos (2021).
Mis ensemble leurs résultats étayent l’hypothèse que les changements de ces phanères – notamment la perte des écailles – ont permis une colonisation efficace de nouveaux milieux (le fond des mers et des lacs) et ont été un facteur important de la radiation des actinopterygiens.
Comment ils ont établi ces conclusions
Les auteurs ont étudié l’évolution des actinopterygiens en se basant sur des arbres phylogénétiques établis en 2018 par des techniques bioinformatiques (ce que l’école appelle la biologie numérique). Ils ont cherché à voir les liens entre taxons, types de couverture tégumentaire et écologie.
Ils se sont basés sur deux phylogénies : l’une plus fine mais moins exhaustive (Hughes et al. 2018) comparant 1105 exons d’un même gène (Cfap58) présent chez toutes les lignées actinoptérygiennes (305 orthologues), et une autre (Rabosky et al. 2018) élaborée sur la base d’un alignement de 27 gènes présents chez 11’638 espèces, incluant près de la moitié des espèces actinoptérygiennes.
Juan Montoya-Burgos précise leurs méthodes « Pour comprendre l’évolution de la couverture tégumentaire et en particulier la transition entre les différentes structures, nous avons étudié les événements de perte d’écailles au cours de l’évolution des actinoptérygiens et abordé la relation fonctionnelle entre le phénotype sans écailles et l’écologie des poissons. De plus, nous avons examiné si l’émergence des plaques osseuses du tronc dépendait de la présence ou de l’absence d’écailles. À cette fin, nous avons utilisé deux phylogénies des actinoptérygiens récemment publiées, l’une comprenant plus de 11 600 espèces, et en utilisant la méthode de stochastic mapping et les méthodes bayésiennes, nous avons reconstruit les événements de perte d’écailles et les acquisitions de plaques osseuses sur le tronc. Pour montrer qu’après un événement de perte d’écailles, les poissons ont tendance à changer leur écologie et à adopter un mode de vie benthique, nous avons utilisé des modèles mixtes linéaires (une famille de méthodes statistiques). »
Cet exemple illustre une fois de plus la place de la bioinformatique (la biologie numérique comme on le dit au DIP) dans la biologie actuelle. Et la nécessité pour l’école de prendre en compte ces nouvelles façons de construire des savoirs.
Des données authentiques disponibles en classe
Juan Montoya‐Burgos a sélectionné pour le projet Jump-To-Science des séquences ADN du gène Cfap58 afin de pouvoir refaire l’alignement avec les élèves. Le gène Cfap58 est un gène qui est présent à une seule copie chez toutes les espèces (single copy gene), qui montre suffisamment de variation et qui n’est pas trop difficile à aligner.
- Un fichier texte ici qui contient les séquences ADN du gène Cfap58 de quelques poissons et du requin (groupe externe), au format FASTA (le code présent après le nom des espèces correspond au numéro d’accession dans la banque de donnée GenBank)
- Un fichier PDF montrant l’arbre phylogénétique que l’on obtient avec ce gène ici
Le chercheur relève que le gène Cfap58 a évolué plus rapidement chez les espèces Hippocampe et PoissonGlobe évoluent plus vite pour ce gène que chez les autres espèces).ici
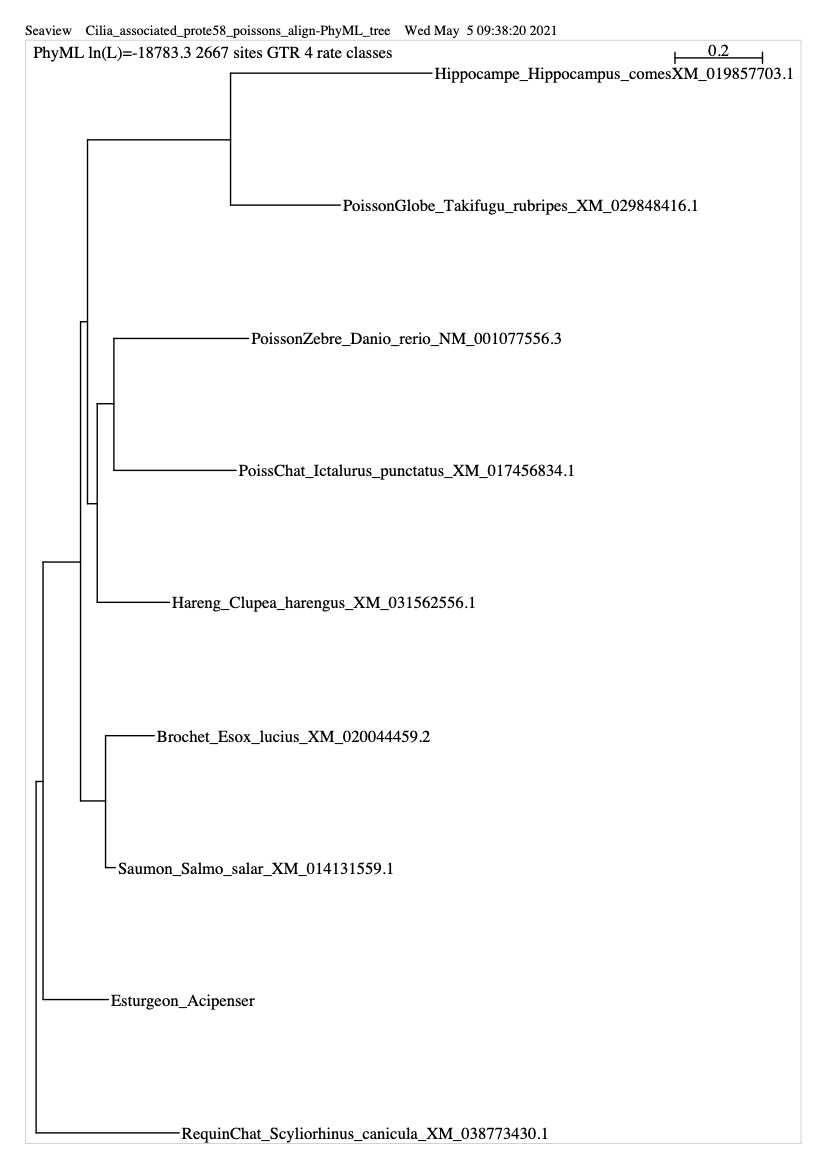{kind=link}
Apprendre les conclusions de la science ce n’est pas faire de la science (Kuhn, 1962)
Eprouver en classe comment on construit une phylogénie par l’alignement des séquences est possible avec les élèves !
- On peut effectuer une versions simplifiée des méthodes des chercheurs : aligner sur UniProt des séquences d’une partie de ces poissons pour observer le grand degré de similarité (visible dans l’image ci-contre) et ici
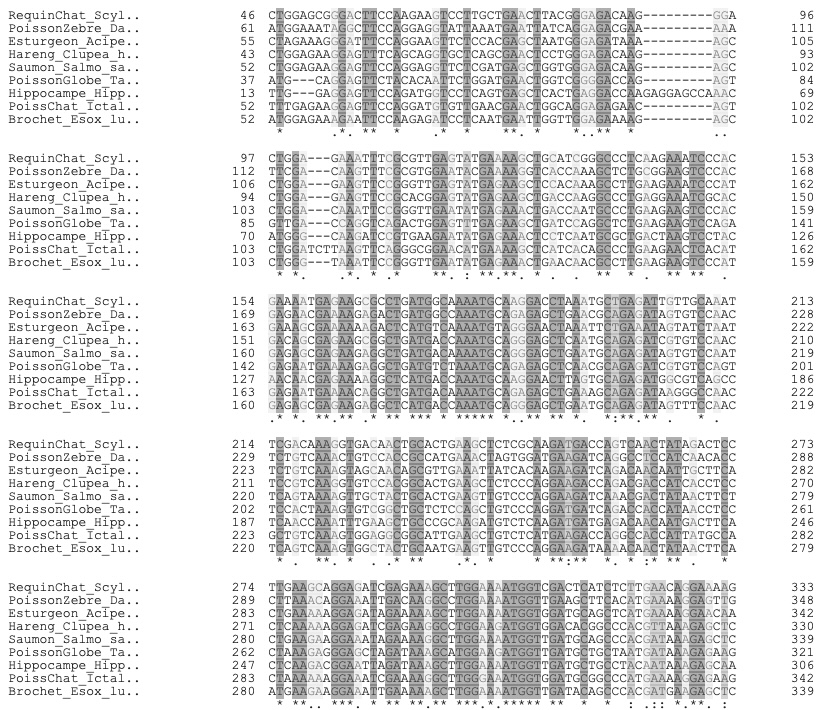{kind=link}
Des lectures pour aller plus loin 
Les rôles et les types d’écailles :
Review sur l’évolution des écailles:
- Sire, J.-Y., Donoghue, P. C. J., & Vickaryous, M. K. (2009). Origin and evolution of the integumentary skeleton in non-tetrapod vertebrates. Journal of Anatomy, 214(4), 409‑440. https://doi.org/10.1111/j.1469-7580.2009.01046.x
Les écailles, de la biologie aux réparations d’os, de cornée :
-
- Zylberberg, L. (2018). The elasmoid scales of teleosts : From structure to bioinspired materials. https://doi.org/10.26028/CYBIUM/2018-421-001
« Mais alors si il n’a pas d’écailles ça peut pas être un poisson, m’dame ?! »
En effet dans l’enseignement on définit souvent les classes par la présence d’écailles (entre autres), c’est-à dire des critères discriminants comme ceux qu’on emploie dans une clé de détermination. Ce faisant on met surtout en évidence et on augmente les différences, cependant on risque que les élèves ne réalisent pas tout ce qui est en commun dans le taxon. Or la phylogénie s’est construite dans le passé par les caractères morpho-anatomiques communs récents (dérivés) et actuellement plutôt par des modèles d’évolution des séquences d’ADN ou de protéines qui reflètent aussi les liens de parentés (ou l’ascendance commune).
Là ou le biologiste voit l’extraordinaire similitude fondamentale dans le vivant, les élèves ont de la peine à voir plus loin qu’un kaléidoscope d’espèces dont la différence émerveille, mais la structure commune n’est pas perçue. Le discours sur la biodiversité renforce probablement cette idée.
C’est un problème difficile qui a été discuté ici Tous pareils tous différents : un slogan ou des explorations en classe ? L’article de Lemopoulos & Montoya‐Burgos apporte un éclairage actualisé sur ces questions et fournit des données pour la discuter en classe.
Références:
- Lemopoulos, A., & Montoya‐Burgos, J. I. (2021). From scales to armor : Scale losses and trunk bony plate gains in ray‐finned fishes. Evolution Letters, evl3.219. https://doi.org/10.1002/evl3.219
- Hughes, L. C., Ortí, G., Huang, Y., Sun, Y., Baldwin, C. C., Thompson, A. W., Arcila, D., Betancur-R, R., Li, C., & Becker, L. (2018). Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proceedings of the National Academy of Sciences, 115(24), 6249‑6254.
- Rabosky, D. L., Chang, J., Title, P. O., Cowman, P. F., Sallan, L., Friedman, M., Kaschner, K., Garilao, C., Near, T. J., & Coll, M. (2018). An inverse latitudinal gradient in speciation rate for marine fishes. Nature, 559(7714), 392‑395.
Remerciements
Ce texte a été co-écrit par Montoya‐Burgos, J. et Lombard, F.avec la précieuse aide de Marie-Claude Blatter pour les aspects Bio-Informatique