
Un espoir contre la malaria identifié grâce aux bases de données et aux simulations
Le parasite de la malaria s’est particulièrement bien adapté au cours de son évolution, car il est équipé pour développer des résistances aux traitements. Le manque de nouvelles approches thérapeutiques contribue également à la persistance de ce fléau d’envergure mondiale. Une étude conduite par Didier Picard, professeur à la Faculté des sciences de l’Université de Genève (UNIGE), décrit une nouvelle classe de molécules ciblant les deux problèmes à la fois. A l’aide d’outils informatiques de modélisation ultra sophistiqués, les chercheurs sont parvenus à identifier un type de molécules candidates toxiques pour le pathogène, mais pas pour les globules rouges humains infectés. L’étude, menée en collaboration avec des chercheurs de l’Ecole de Pharmacie Genève-Lausanne (EPGL) et de l’Université de Bâle, est publiée dans le Journal of Medicinal Chemistry. L’article est ici (les membres Expériment@l peuvent obtenir ces articles… cette publication est un tremplin vers les articles de recherche )
La malaria tue annuellement près d’un million de personnes, surtout des enfants
La malaria était encore courante autour de la méditerranée jusqu’à vers 1950 selon Hedrick, P. W. (2011). Elle frappe 243 million de personnes (2008) et on estime à 863 000 les morts causés annuellement par la malaria (WHO, 2009), soit un enfant toutes les 40 secondes et plus de 2000 par jour (Sachs, J., & al. ,2002) . La plus grande partie de cette mortalité est due à Plasmodium falciparum. Selon Hedrick, P. W. (2011) elle constitue la pression de sélection la plus forte dans l’histoire évolutive humaine récente. (les membres Expériment@l peuvent obtenir ces articles…)
« The high mortality and widespread impact of malaria have resulted in this disease being the strongest evolutionary selective force in recent human history, and genes that confer resistance to malaria provide some of the best-known case studies of strong positive selection in modern humans.« Hedrick, P. W. (2011).
On pense naturellement à l’anémie falciforme (drépanocytose) exemple classique dans les écoles, mais Hedrick, P. W. (2011) discutent aussi dans leur article Population genetics of malaria resistance in humans : de la déficience en G6PD, Duffy, ovalocytose, les variantes ABO et HLA (human leukocyte antigen).
La recherche de thérapies contre la malaria se heurte à divers obstacles, dont la variabilité du parasite, son cycle complexe, (cf. par exemple Winzeler, E. A. (2008). Malaria research in the post-genomic era), mais aussi à son métabolisme assez particulier et très bien adapté aux globules rouges où une partie de son cycle se déroule comme nous le verrons plus bas.
Les protéines chaperonnes protègent des effets du chaud
Un des symptômes de la malaria est des fièvres très intenses (43°C) et récurrentes tous les 2-3 jours. Cette température élevée semble favoriser le parasite. A cette température les protéines peuvent changer de conformation, et on sait que les protéines Hsp (Heat shock protein) – induites à ces températures – sont des chaperonnes qui guident la conformation des protéines. Selon Acharya, P.,et al (2007), le parasite exploite ces protéines Hsp pour détourner et réguler le fonctionnement des protéines du globule rouge (érythrocyte). On se souvient qu’il n’a pas de noyau et que la production de nouvelles protéines n’y est pas possible.
« Even though mature human erythrocytes present neither an active metabolism nor transport apparatus, the parasite spends a significant portion of its life cycle in them. While this may be an efficient immune-evasion strategy, the parasites must compensate for the lack of biochemical support in the host. The parasite needs to re-establish new transport mechanisms, develop elaborate secretory apparatus, and recruit available host resources most efficiently. Studies performed in various laboratories suggest that the parasite has evolved to rely on its chaperone systems as a combined strategy to support re-structuring of the host cell as well as respond to stressful environment of the host. » Acharya, P.,et al (2007). (les membres Expériment@l peuvent obtenir ces articles…)
Les chaperonnes déforment et détournent les fonctions de protéines humaines pour permettre le cycle du parasite
Il semble (Acharya, P.,et al. 2007) qu’une complexe restructuration des protéines de la cellule-hôte permette au parasite son développement et de résister aux conditions difficiles de son environnement. En cela le plasmodium diffère d’autres parasites intracellulaires comme les virus par exemple qui contrôlent la production des protéines, puisqu’il n’y a pas de biosynthèse des protéines dans les érythrocytes.
Le modèle simplifié pour comprendre les protéines présentes dans une cellule et leur activité (« dogme central de la biologie ») posait que la structure des protéines est déterminées par leur séquence qui est déterminée par l’ARN et l’ADN. Ici à partir de protéines avec la même séquence, le parasite arriverait à les modifier pour obtenir des fonctions nécessaires à son cycle biologique en activant des protéines chaperonnes qui les déforment !
Des cibles potentielles
Parce qu’elles sont cruciales pour le parasite, ces protéines Hsp constituent donc une cible thérapeutiques potentielle. L’une de ces protéines (Hsp90) est étudiée depuis longtemps par l’équipe du Pr. Picard à l’UniGE. Or la séquence de cette protéine est très conservée (on la retrouve très semblable chez des organismes très différents; on parle d’orthologue) ce qui est en général interprété comme une pression de sélection forte: les organismes que le hasard a muni de variantes mutées différentes ne se développent pas ou en tous cas se reproduisent moins. Vous et moi sommes donc les descendants de ceux qui ont hérité de gènes pour Hsp fonctionnels et comme il y a peu de conformations fonctionnelles possibles nous avons probablement des Hsp très similaires.
Du coup les protéines Hsp90 sont aussi très semblables entre diverses espèces notamment entre P. falciparum et H. sapiens. Dans l’idéal, on cherche à interférer avec une protéine qui est très peu variable et cruciale pour le pathogène, mais très différente de celles des humains. On ne peut donc pas facilement inhiber Hsp90 sans atteindre aussi nos cellules.
« The high sequence conservation of druggable pockets of closely related proteins can make it challenging to develop selective inhibitors. » Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014) (les membres Expériment@l peuvent obtenir ces articles…)
C’est là que l’étude menée par le Pr. Picard apporte des informations critiques : Ils ont observé que malgré une séquence très très semblable de PfHsp90 (Hsp90 deP. falciparum ) et Hsp90 (humaine) leur conformation dynamique est différente (cf figure 2). Le doctorant Tai Wang a notamment utilisé des outils informatiques de modélisation des protéines ultra sophistiqués (cf Le prix Nobel de chimie : un changement du paradigme de recherche ? ) pour caractériser les diverses conformations tri-dimensionnelle de l’HSP90 parasitaire.

Fig 2: En jaune la partie qui pourrait lier une molécule thérapeutique. En rouge et vert les séquences et les domaines de protéine délimitant la PfHsp90-specific hydrophobic pocket étudiée [img] source Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014)
Encore de la science in silico
Dans le labo du Pr Picard, il a découvert une niche capable de lier des substances inhibitrices, totalement absente chez son alter ego humain.
Ce n’est pas tant la séquence que la structure dynamique qui diffère.
« Upon examining the available crystal structures in detail, we discovered an area of structural divergence in PfHsp90 that arises without underlying sequence divergence. […] Thus, our structural and computational discoveries led us to conclude that the minor primary sequence differences between the human and Plasmodium Hsp90s translate into far more important structural differences » (les membres Expériment@l peuvent obtenir ces articles…)
Screening
A l’aide d’un superordinateur, le chercheur a ensuite effectué le criblage d’une librairie virtuelle contenant plus d’un million de composés chimiques, en retenant ceux pouvant se loger dans cette niche. Ce criblage in silico l’a mené à sélectionner cinq candidats, des 7-azaindole .

Fig 3 : Représentation modélisée d’une molécule inhibitrice ( jaune) dans une ‘niche’ de la protéine HSP90 du parasite (rose). [img] source Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014)
Simulation en temps réel
Ces expériences ont ensuite été complétées par une technique de modélisation ‘en temps réel’. «Les simulations m’ont permis d’analyser la dynamique d’interaction entre l’HSP90 et les candidats et de découvrir des inhibiteurs qui interagissent de façon spécifique avec la chaperonne du Plasmodium falciparum».

Fig 4: L’ATP-binding pocket de PfHsp90 a une extension spécifique du Plasmodium, malgré une séquence identique à la Hsp90 humaine mais diffère par la structure tertiaire et sa dynamique. En rouge et vert les séquences et les domaines de protéine délimitant la PfHsp90-specific hydrophobic pocket étudiée [img] source Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014)
Vérification in vivo
Les molécules ont ensuite été testées in vitro dans différents systèmes. Les biologistes ont notamment démontré la toxicité de ces inhibiteurs sur des cultures de Plasmodium falciparum, à des doses capables de tuer les parasites sans affecter les globules rouges infectés. «Ces molécules, récemment brevetées, font partie d’un groupe de composés apparentés aux 7-azaindoles, qui se lient exclusivement à l’HSP90 du parasite, mais pas à la forme humaine. La prochaine étape sera de les peaufiner pour pouvoir effectuer les tests cliniques», conclut Didier Picard.
mais le plus intéressant est la méthode qui combine biologie in silico, simulation chimiques et vérification in vivo.
« The strength of our approach is that it takes advantage of conformational differences that may be apparent in static crystal structure, but even more so upon comparing dynamic properties. It is not that human Hsp90s, and the IXXSG mutant of PfHsp90 for that matter, cannot at all adopt the conformation that is favored by PfHsp90. They can, but ligand binding is entropically penalized when the GHL conformation is not constrained. As a result, the species selectivity of our current compounds is primarily due to a thermodynamic difference in binding to PfHsp90 compared to human Hsp90s. « Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014) (les membres Expériment@l peuvent obtenir ces articles…)
Et plus loin
« Our discovery of a promising new pharmacophore constitutes a proof of concept for our novel workflow. The detailed scrutiny of the target protein allowed us to set up a large-scale in silico screen for compounds that could take advantage of a Pf-specific hydrophobic pocket. The analysis of the differences in conformational dynamics done in parallel set the stage for predicting the binding site and explaining the species selectivity of compounds that came out of the VS and experimental pipeline. Demonstrated here for two orthologous proteins, this concept may be more generally applicable, notably to closely related proteins of the same species. In drug screens, conformationally constrained mutants could help to eliminate compounds that target closely related unwanted targets » Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014) (les membres Expériment@l peuvent obtenir ces articles…)
Une nouvelle façon de faire de la recherche… de nouvelles possibilités d’enseigner ?
La complémentarité entre les approches in silico et in vitro dans cette étude manifeste bien le rôle crucial que la biologie et la chimie de l’information prennent dans la recherche actuelle. L’expérimentation n’est pas seulement affaire de molécules et de manipulation de pipettes, mais peut-être d’abord le traitement d’information pour explorer les hypothèses les confronter les tester… En quoi cela change-t-il ce qui se passe dans les labos et TP dans lesquels nos élèves apprennent ce qu’est la chimie et la biologie ?
 L’activité du chercheur qui produit un savoir digne d’être publié est ici principalement la manipulation de modèles pour traiter les informations des bases de données et de simulations derrière un ordinateur.
Le prix Nobel de Chimie 2014 a ouvert une voie qui a changé la façon de faire la chimie, l’avènement des bases de données de génomes que le projet génome humain a révélé au public une nouvelle façon de faire la biologie, tous deux manifestent une transformation en profondeur de nos sciences.
L’activité du chercheur qui produit un savoir digne d’être publié est ici principalement la manipulation de modèles pour traiter les informations des bases de données et de simulations derrière un ordinateur.
Le prix Nobel de Chimie 2014 a ouvert une voie qui a changé la façon de faire la chimie, l’avènement des bases de données de génomes que le projet génome humain a révélé au public une nouvelle façon de faire la biologie, tous deux manifestent une transformation en profondeur de nos sciences.
Bien sûr à la fin on a vérification dans le wet lab, pipette à la main sur des cellules et des être vivants… Ce qui justifie la publication c’est tout le traitement de modèles sur des extraits de banques de données : cette expérimentation-là… pas la manipulation qui confirme.
Ainsi ce qui définit le biologie et le chimiste n’est peut-être pas tant des gestes que le type d’information qu’il traite…
Pour comprendre ces expériences, que doit savoir un enseignant ?
Quelles possibilités en classe offrent les accès aisés aux données authentiques (les tablettes sont presque en classe… d’interdites elles vont devenir obligatoires et prescrites d’ici peu) ?
Vos élèves peuvent par exemple aller voir
- La structure 3-D de PfHsp90 sur PDB
- La séquence de la protéine Hsp90 alpha sur UniProt
- » Molecular chaperone that promotes the maturation, structural maintenance and proper regulation of specific target proteins involved for instance in cell cycle control and signal transduction. Undergoes a functional cycle that is linked to its ATPase activity. This cycle probably induces conformational changes in the client proteins, thereby causing their activation. Interacts dynamically with various co-chaperones that modulate its substrate recognition, ATPase cycle and chaperone function. »
Pour que l’élève comprenne la biologie et la chimie du XXième que doit-on lui faire faire ?
S’agit-il que l’enseignant les montre au projecteur, ou de guider les élèves pour les trouver, ou encore de leur faire comparer Hsp90 ( P07900) et PfHsp90 ( Q8IL32) par un alignement puis discuter les raisons possibles de cette incroyable similitude ?
Il n’y a sûrement pas une réponse unique pour toutes les situations d’enseignement.
Il y a toute une didactique à inventer… et déjà de magnifiques opportunités de permettre aux élèves d’expérimenter en classe ?
Est-il possible de l’ignorer complètement ?
La malaria tue annuellement près d’un million de personnes, surtout des enfants
La malaria était encore courante autour de la méditerranée jusqu’à vers 1950 selon Hedrick, P. W. (2011). Elle frappe 243 million de personnes (2008) et on estime à 863 000 les morts causés annuellement par la malaria (WHO, 2009), soit un enfant toutes les 40 secondes et plus de 2000 par jour (Sachs, J., & al. ,2002) . La plus grande partie de cette mortalité est due à Plasmodium falciparum. Selon Hedrick, P. W. (2011) elle constitue la pression de sélection la plus forte dans l’histoire évolutive humaine récente. (les membres Expériment@l peuvent obtenir ces articles…)
« The high mortality and widespread impact of malaria have resulted in this disease being the strongest evolutionary selective force in recent human history, and genes that confer resistance to malaria provide some of the best-known case studies of strong positive selection in modern humans.« Hedrick, P. W. (2011).
On pense naturellement à l’anémie falciforme (drépanocytose) exemple classique dans les écoles, mais Hedrick, P. W. (2011) discutent aussi dans leur article Population genetics of malaria resistance in humans : de la déficience en G6PD, Duffy, ovalocytose, les variantes ABO et HLA (human leukocyte antigen).
La recherche de thérapies contre la malaria se heurte à divers obstacles, dont la variabilité du parasite, son cycle complexe, (cf. par exemple Winzeler, E. A. (2008). Malaria research in the post-genomic era), mais aussi à son métabolisme assez particulier et très bien adapté aux globules rouges où une partie de son cycle se déroule comme nous le verrons plus bas.

Les protéines chaperonnes protègent des effets du chaud
Un des symptômes de la malaria est des fièvres très intenses (43°C) et récurrentes tous les 2-3 jours. Cette température élevée semble favoriser le parasite. A cette température les protéines peuvent changer de conformation, et on sait que les protéines Hsp (Heat shock protein) – induites à ces températures – sont des chaperonnes qui guident la conformation des protéines. Selon Acharya, P.,et al (2007), le parasite exploite ces protéines Hsp pour détourner et réguler le fonctionnement des protéines du globule rouge (érythrocyte). On se souvient qu’il n’a pas de noyau et que la production de nouvelles protéines n’y est pas possible.
« Even though mature human erythrocytes present neither an active metabolism nor transport apparatus, the parasite spends a significant portion of its life cycle in them. While this may be an efficient immune-evasion strategy, the parasites must compensate for the lack of biochemical support in the host. The parasite needs to re-establish new transport mechanisms, develop elaborate secretory apparatus, and recruit available host resources most efficiently. Studies performed in various laboratories suggest that the parasite has evolved to rely on its chaperone systems as a combined strategy to support re-structuring of the host cell as well as respond to stressful environment of the host. » Acharya, P.,et al (2007). (les membres Expériment@l peuvent obtenir ces articles…)
Les chaperonnes déforment et détournent les fonctions de protéines humaines pour permettre le cycle du parasite
Il semble (Acharya, P.,et al. 2007) qu’une complexe restructuration des protéines de la cellule-hôte permette au parasite son développement et de résister aux conditions difficiles de son environnement. En cela le plasmodium diffère d’autres parasites intracellulaires comme les virus par exemple qui contrôlent la production des protéines, puisqu’il n’y a pas de biosynthèse des protéines dans les érythrocytes.
Le modèle simplifié pour comprendre les protéines présentes dans une cellule et leur activité (« dogme central de la biologie ») posait que la structure des protéines est déterminées par leur séquence qui est déterminée par l’ARN et l’ADN. Ici à partir de protéines avec la même séquence, le parasite arriverait à les modifier pour obtenir des fonctions nécessaires à son cycle biologique en activant des protéines chaperonnes qui les déforment !
Des cibles potentielles
Parce qu’elles sont cruciales pour le parasite, ces protéines Hsp constituent donc une cible thérapeutiques potentielle. L’une de ces protéines (Hsp90) est étudiée depuis longtemps par l’équipe du Pr. Picard à l’UniGE. Or la séquence de cette protéine est très conservée (on la retrouve très semblable chez des organismes très différents; on parle d’orthologue) ce qui est en général interprété comme une pression de sélection forte: les organismes que le hasard a muni de variantes mutées différentes ne se développent pas ou en tous cas se reproduisent moins. Vous et moi sommes donc les descendants de ceux qui ont hérité de gènes pour Hsp fonctionnels et comme il y a peu de conformations fonctionnelles possibles nous avons probablement des Hsp très similaires.
Du coup les protéines Hsp90 sont aussi très semblables entre diverses espèces notamment entre P. falciparum et H. sapiens. Dans l’idéal, on cherche à interférer avec une protéine qui est très peu variable et cruciale pour le pathogène, mais très différente de celles des humains. On ne peut donc pas facilement inhiber Hsp90 sans atteindre aussi nos cellules.
« The high sequence conservation of druggable pockets of closely related proteins can make it challenging to develop selective inhibitors. » Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014) (les membres Expériment@l peuvent obtenir ces articles…)
C’est là que l’étude menée par le Pr. Picard apporte des informations critiques : Ils ont observé que malgré une séquence très très semblable de PfHsp90 (Hsp90 deP. falciparum ) et Hsp90 (humaine) leur conformation dynamique est différente (cf figure 2). Le doctorant Tai Wang a notamment utilisé des outils informatiques de modélisation des protéines ultra sophistiqués (cf Le prix Nobel de chimie : un changement du paradigme de recherche ? ) pour caractériser les diverses conformations tri-dimensionnelle de l’HSP90 parasitaire.
Fig 2: En jaune la partie qui pourrait lier une molécule thérapeutique. En rouge et vert les séquences et les domaines de protéine délimitant la PfHsp90-specific hydrophobic pocket étudiée [img] source Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014)
Encore de la science in silico
Dans le labo du Pr Picard, il a découvert une niche capable de lier des substances inhibitrices, totalement absente chez son alter ego humain.
Ce n’est pas tant la séquence que la structure dynamique qui diffère.
« Upon examining the available crystal structures in detail, we discovered an area of structural divergence in PfHsp90 that arises without underlying sequence divergence. […] Thus, our structural and computational discoveries led us to conclude that the minor primary sequence differences between the human and Plasmodium Hsp90s translate into far more important structural differences » (les membres Expériment@l peuvent obtenir ces articles…)
Screening
A l’aide d’un superordinateur, le chercheur a ensuite effectué le criblage d’une librairie virtuelle contenant plus d’un million de composés chimiques, en retenant ceux pouvant se loger dans cette niche. Ce criblage in silico l’a mené à sélectionner cinq candidats, des 7-azaindole .
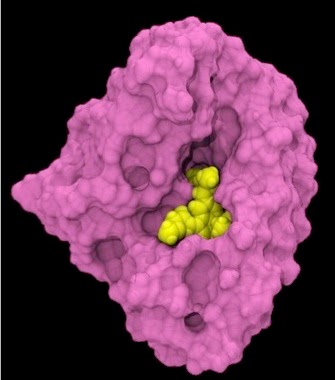
Fig 3 : Représentation modélisée d’une molécule inhibitrice ( jaune) dans une ‘niche’ de la protéine HSP90 du parasite (rose). [img] source Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014)
Simulation en temps réel
Ces expériences ont ensuite été complétées par une technique de modélisation ‘en temps réel’. «Les simulations m’ont permis d’analyser la dynamique d’interaction entre l’HSP90 et les candidats et de découvrir des inhibiteurs qui interagissent de façon spécifique avec la chaperonne du Plasmodium falciparum».
Fig 4: L’ATP-binding pocket de PfHsp90 a une extension spécifique du Plasmodium, malgré une séquence identique à la Hsp90 humaine mais diffère par la structure tertiaire et sa dynamique. En rouge et vert les séquences et les domaines de protéine délimitant la PfHsp90-specific hydrophobic pocket étudiée [img] source Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014)
Vérification in vivo
Les molécules ont ensuite été testées in vitro dans différents systèmes. Les biologistes ont notamment démontré la toxicité de ces inhibiteurs sur des cultures de Plasmodium falciparum, à des doses capables de tuer les parasites sans affecter les globules rouges infectés. «Ces molécules, récemment brevetées, font partie d’un groupe de composés apparentés aux 7-azaindoles, qui se lient exclusivement à l’HSP90 du parasite, mais pas à la forme humaine. La prochaine étape sera de les peaufiner pour pouvoir effectuer les tests cliniques», conclut Didier Picard.
mais le plus intéressant est la méthode qui combine biologie in silico, simulation chimiques et vérification in vivo.
« The strength of our approach is that it takes advantage of conformational differences that may be apparent in static crystal structure, but even more so upon comparing dynamic properties. It is not that human Hsp90s, and the IXXSG mutant of PfHsp90 for that matter, cannot at all adopt the conformation that is favored by PfHsp90. They can, but ligand binding is entropically penalized when the GHL conformation is not constrained. As a result, the species selectivity of our current compounds is primarily due to a thermodynamic difference in binding to PfHsp90 compared to human Hsp90s. « Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014) (les membres Expériment@l peuvent obtenir ces articles…)
Et plus loin
« Our discovery of a promising new pharmacophore constitutes a proof of concept for our novel workflow. The detailed scrutiny of the target protein allowed us to set up a large-scale in silico screen for compounds that could take advantage of a Pf-specific hydrophobic pocket. The analysis of the differences in conformational dynamics done in parallel set the stage for predicting the binding site and explaining the species selectivity of compounds that came out of the VS and experimental pipeline. Demonstrated here for two orthologous proteins, this concept may be more generally applicable, notably to closely related proteins of the same species. In drug screens, conformationally constrained mutants could help to eliminate compounds that target closely related unwanted targets » Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014) (les membres Expériment@l peuvent obtenir ces articles…)
Une nouvelle façon de faire de la recherche… de nouvelles possibilités d’enseigner ?
L’activité du chercheur qui produit un savoir digne d’être publié est ici principalement la manipulation de modèles pour traiter les informations des bases de données et de simulations derrière un ordinateur.Bien sûr à la fin on a vérification dans le wet lab, pipette à la main sur des cellules et des être vivants… Ce qui justifie la publication c’est tout le traitement de modèles sur des extraits de banques de données : cette expérimentation-là… pas la manipulation qui confirme.
Pour comprendre ces expériences, que doit savoir un enseignant ?
- La structure 3-D de PfHsp90 sur PDB
- La séquence de la protéine Hsp90 alpha sur UniProt
- » Molecular chaperone that promotes the maturation, structural maintenance and proper regulation of specific target proteins involved for instance in cell cycle control and signal transduction. Undergoes a functional cycle that is linked to its ATPase activity. This cycle probably induces conformational changes in the client proteins, thereby causing their activation. Interacts dynamically with various co-chaperones that modulate its substrate recognition, ATPase cycle and chaperone function. »
Pour que l’élève comprenne la biologie et la chimie du XXième que doit-on lui faire faire ?
S’agit-il que l’enseignant les montre au projecteur, ou de guider les élèves pour les trouver, ou encore de leur faire comparer Hsp90 ( P07900) et PfHsp90 ( Q8IL32) par un alignement puis discuter les raisons possibles de cette incroyable similitude ?

Il n’y a sûrement pas une réponse unique pour toutes les situations d’enseignement.
Il y a toute une didactique à inventer… et déjà de magnifiques opportunités de permettre aux élèves d’expérimenter en classe ?
Est-il possible de l’ignorer complètement ?
Sources : (les membres Expériment@l peuvent obtenir ces articles…)
- Acharya, P., Kumar, R., & Tatu, U. (2007). Chaperoning a cellular upheaval in malaria: Heat shock proteins in Plasmodium falciparum. Molecular and Biochemical Parasitology, 153(2), 85‑94. doi:10.1016/j.molbiopara.2007.01.009
- Wang, T., Bisson, W. H., Mäser, P., Scapozza, L., & Picard, D. (2014). Differences in Conformational Dynamics between Plasmodium falciparum and Human Hsp90 Orthologues Enable the Structure-Based Discovery of Pathogen-Selective Inhibitors. Journal of Medicinal Chemistry. doi:10.1021/jm401801t
- Hedrick, P. W. (2011). Population genetics of malaria resistance in humans. Heredity, 107(4), 283‑304. doi:10.1038/hdy.2011.16
- Roy, N., Nageshan, R. K., Ranade, S., & Tatu, U. (2012). Heat shock protein 90 from neglected protozoan parasites. Biochimica et Biophysica Acta (BBA) – Molecular Cell Research, 1823(3), 707‑711. doi:10.1016/j.bbamcr.2011.12.003
-
Sachs, J., & Malaney, P. (2002). The economic and social burden of malaria. Nature, 415(6872), 680‑685. doi:10.1038/415680a
- Winzeler, E. A. (2008). Malaria research in the post-genomic era. Nature, 455(7214), 751‑756. doi:10.1038/nature07361
- Labie, D. (2005). La fièvre récurrente du paludisme : un atout majeur pour le parasite. Erudit.org: médecine sciences, 21(3), 243‑244.
- http://pubs.acs.org/doi/pdf/10.1021/jm401801t
La plateforme Expériment@l vous offre l’accès a ces articles Comment s’inscrire à Expériment@l / Comment Obtenir un article mentionné : Get-a-doi